ARTÍCULO DE REVISIÓN
Helmintosis y autoinmunidad
Helminth infections and autoimmunity
Dr. Luis Fonte Galindo , Dra. Gissel García Menéndez, Lic. Jessica Baldriche Acosta , Dra. Annia Fong González, Dra. Yuliet Méndez Sutil
Instituto de Medicina Tropical "Pedro Kourí". La Habana, Cuba.
RESUMEN
Las evidencias epidemiológicas, clínicas e inmunológicas de estudios en humanos y los datos obtenidos de experimentos en modelos animales ofrecen un soporte creciente al criterio de que las infecciones por helmintos tienen un efecto protector contra entidades patológicas que transcurren con desregulación del sistema inmunitario, tales como enfermedades autoinmunes y algunas alteraciones inflamatorias idiopáticas. A partir de este precedente, el objetivo de este trabajo fue revisar y analizar lo publicado sobre helmintosis, regulación de las respuestas inmunitarias y eventos autoinmunes e inflamatorios. Los análisis realizados permiten concluir que la regulación de las respuestas inmunitarias del hospedero por los helmintos repercute en la frecuencia e intensidad de eventos autoinmunes e inflamatorios. En aras de una práctica médica de mejor calidad, las consecuencias clínicas y terapéuticas de esas repercusiones deben ser conocidas por los profesionales relacionados con el diagnóstico y tratamiento de enfermedades autoinmunes y alteraciones inflamatorias idiopáticas.
Palabras clave: helmintosis, helmintos, inmunorregulación, autoinmunidad, inflamación.
ABSTRACT
Epidemiological, clinical and immunological evidence from human studies and data obtained from experiments in animal models offer increased support to the view that helminth infections have a protective effect against pathological entities that run with deregulation of the immune system, such as illness idiopathic autoimmune and inflammatory changes some. From this precedent, the objective of this study was to review and analyze the literature on helminth infections, regulation of immune responses and autoimmune and inflammatory events. Studies support the conclusion that regulation of immune responses by helminth hosts affects frequency and intensity of autoimmune and inflammatory events. In order to better quality medical practice, clinical and therapeutic implications of these impacts should be known by professionals in diagnosis and treatment of idiopathic autoimmune diseases and inflammatory disorders.
Keywords : helminthes infections, helminths, immunoregulation, autoimmunity, inflammation.
INTRODUCCIÓN
Durante las últimas cinco décadas, se ha producido un incremento en la incidencia de enfermedades autoinmunes y alteraciones inflamatorias idiopáticas en los países económicamente desarrollados (fundamentalmente, en Estados Unidos y Europa) y, más recientemente, en las áreas urbanizadas de las naciones en desarrollo1-2. Ese incremento ha sido tan acelerado que no podría ser explicado por factores genéticos (sin que ello niegue la existencia de predisposición genética a la mayoría de esas entidades). En consecuencia, se considera que factores ambientales serían protagónicos en la emergencia de esas enfermedades1-7. Datos epidemiológicos parecen proporcionar suficiente soporte al postulado de que las infecciones por helmintos podrían proveer alguna protección contra eventos autoinmunes e inflamatorios 2.
Un corolario de ese enunciado sería que las estrategias de higienización, saneamiento ambiental y desparasitación en las áreas urbanizadas hacen a las personas que en ellas viven más vulnerables al desarrollo de enfermedades autoinmunes y alteraciones inflamatorias idiopáticas2,8. Aunque la mayoría de los datos epidemiológicos y un cúmulo creciente de resultados experimentales apoyan el mencionado postulado, la comprensión precisa de la relación entre infecciones por helmintos y fenómenos autoinmunes es hoy tema de intensos estudios y punto de partida para prometedoras aproximaciones terapéuticas2,5.
LAS INFECCIONES POR HELMINTOS PROTEGEN CONTRA ENFERMEDADES AUTOINMUNES
Las evidencias en favor de que las infecciones por helmintos protegen contra enfermedades autoinmunes y alteraciones inflamatorias idiopáticas pueden ser consideradas de tres tipos: evidencias epidemiológicas basadas en metanálisis, evidencias epidemiológicas basadas en hallazgos serológicos y evidencias experimentales.
Evidencias epidemiológicas basadas en metanálisis
Los metanálisis de observaciones, registros y publicaciones realizadas en diferentes áreas geográficas, principalmente en Estados Unidos y Europa, favorecen la creencia de que las infecciones por helmintos protegen contra enfermedades autoinmunes y alteraciones inflamatorias idiopáticas. Así:
- En Estados Unidos y Europa, la enfermedad de Crohn emergió primero en las poblaciones más opulentas que vivían en condiciones higiénicas favorables y en latitudes más al norte, donde las temperaturas más frías hacían menos propicios los escenarios para la transmisión de las infecciones por helmintos9.
- En Estados Unidos, el último grupo racial donde la enfermedad de Crohn se hizo presente fue en los individuos de origen africano, quienes, en promedio, son más pobres que las personas de origen europeo9.
- En estudios realizados en varios países, se ha observado una relación negativa entre incidencia de esclerosis múltiple y prevalencia de infección por helmintos10.
- Las enfermedades autoinmunes son más frecuentes entre los individuos que viven en Europa Occidental en relación a los que habitan en Europa del Este9.
- En los remanentes de poblaciones americanas nativas, en los que las infecciones por helmintos son de alta prevalencia, la frecuencia de enfermedades intestinales inflamatorias es baja9.
- Los latinos que llegaron a Estados Unidos después de la adolescencia no desarrollan desórdenes inflamatorios intestinales. Sin embargo, sus hijos nacidos en ese país, donde las condiciones sanitarias son mejores y las posibilidades de infectar por helmintos son mínimas, tienen mayor riesgo de padecer de enfermedades autoinmunes y desórdenes inflamatorios9.
- La distribución mundial de las enfermedades intestinales inflamatorias es prácticamente la imagen especular de las áreas endémicas de infecciones por helmintos2.
Evidencias epidemiológicas basadas en hallazgos serológicos
Aún son escasas las pesquisas que tratan de evidenciar con herramientas serológicas alguna relación entre infecciones por helmintos y fenómenos autoinmunes. Entre ellas, dos merecen ser especialmente mencionadas:
- Un estudio reciente que halló que individuos infectados con esquistosomas tenían menores títulos de autoanticuerpos que la población libre del parásito y que los títulos de esos anticuerpos se incrementaron cuando las personas infectadas fueron tratadas con praziquantel, el medicamento antiparasitario más frecuentemente utilizado en esos casos11.
- Un trabajo realizado en Argentina en pacientes con esclerosis múltiple en remisión que demostró que cuando estos eran infectados por parásitos gastrointestinales (Ascaris lumbricoides, Trichuris trichiura, Strongyloides stercoralis, Hymenolepis nana y Enterobius vermicularis) permanecían controlados durante más tiempo y cuando ocurrían recrudecimientos de la enfermedad estos eran más leves. Significativamente, los pacientes con supresión de la enfermedad asociada con la infección por helmintos mostraban niveles más elevados de las citoquinas reguladoras IL-10 y TGF-β que los individuos no infectados12-13.
Evidencias experimentales
Las enfermedades autoinmunes son considerablemente menos prevalentes que las alérgicas y, consecuencia de ello, son relativamente escasas las investigaciones epidemiológicas que han podido realizarse para conocer el efecto de las infecciones por helmintos sobre esos desórdenes 5. Para llenar ese vacío, numerosos trabajos experimentales han sido realizados. A continuación se mencionan algunos de los resultados más trascendentes:
- En fecha tan temprana como 1975, Pearson y Taylor reportaron que en ratas en las que se habían inducido cambios artríticos mediante la administración de adyuvante completo de Freund, la infección por el nematodo Symphacia obvelataatenuaba esas manifestaciones14.
- En la artritis inducida por colágeno tipo II, la infección por Schistosoma mansonireduce la producción de anticuerpos contra colágeno y de citoquinas inflamatorias, a la vez que los daños histopatológicos correlacionan negativamente con la carga parasitaria 15.
- La infección por S.mansoni inhibe el desarrollo de diabetes tipo I en ratones NOD (del inglés non obese diabetic), ratones que desarrollan diabetes espontáneamente después de los 6 meses de edad16.
- El desarrollo de lesiones en el modelo murino de esclerosis múltiple puede ser inhibido por la infección por varios parásitos, como fue primeramente demostrado con S.mansoni17-18.
- Las enfermedades inflamatorias intestinales, entre ellas la colitis ulcerativa y la enfermedad de Crohn, son desórdenes frecuentes, y en ocasiones graves, del humano19. Varios modelos de estas enfermedades, generalmente diferenciados por su agente inductor, han sido desarrollados en ratones. El desarrollo de colitis inducida por ácido dinitrobenzeno sulfónico puede ser atenuado por productos de Hymenolepis diminuta. En otro estudio fue demostrado que los efectos supresores de H. diminuta eran dependientes de la acción de macrófagos alternativamente activados y que incluso la administración de estos macrófagos en ratones vírgenes impedía la inducción de la colitis20. En correspondencia con este hallazgo, es muy interesante la observación de que la remisión de la enfermedad de Crohn en los humanos está asociada con la acumulación de macrófagos alternativamente activados en la pared intestinal5,21.
MECANISMOS DE PROTECCIÓN CONTRA ENFERMEDADES AUTOINMUNES E INFLAMATORIAS
Los resultados hoy disponibles permiten concluir que esa protección contra enfermedades autoinmunes e inflamatorias es consecuencia de dos fenómenos inducidos por estos parásitos: la regulación de las respuestas inmunitarias del hospederoy cambios en la flora bacteriana de este 2-3,5,7,9,22-28.
Regulación de las respuestas inmunitarias del hospedero
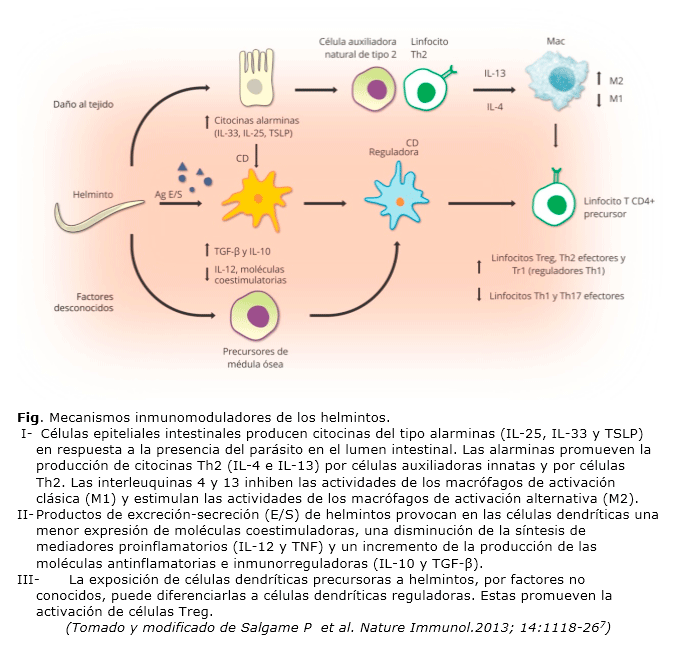
Los helmintos interfieren en las respuestas inmunitarias de sus respectivos hospederos debido a una compleja madeja de mecanismos regulatorios 29-34. Estos, por las secuencias en que tienen lugar, pueden agruparse en tres grandes ejes (sin que ello excluya la posibilidad de interacciones entre componentes de las tres vertientes) (Fig.). A continuación se describen los eventos más trascendentales que tienen lugar en los mencionados ejes regulatorios.
Producción de citoquinas del tipo alarminas (IL-25, IL-33 y TSLP) por células epiteliales intestinales en presencia del parásito .
- Las células epiteliales intestinales (y posiblemente otras células en otros órganos y tejidos) producen IL-25, IL-33 y TSLP (del inglés thimic stromal limphopoietin) en respuesta a la presencia del parásito en el lumen intestinal26,35. Las IL-25 e IL-33, como se verá más adelante, están involucradas en el estímulo a la producción primaria de mediadores tipo 2. En cambio, TSLP parece estar más relacionado con la limitación de la producción de IL-12 por células dendríticas36-37.
- Las células CD4+ Th2, protagónicas en las respuestas adaptativas a la infección por helmintos, son las mayores fuentes de citoquinas tipo 238. Sin embargo, estudios de la última década han identificado una nueva línea de células innatas en los ganglios linfáticos mesentéricos que producen grandes cantidades de IL-4, IL-5 e IL-1339-42. Estas células, frecuentemente denominadas células auxiliadoras tipo 2 innatas42, son respondedoras a las alarminas IL-25 e IL-33 secretadas por las células epiteliales intestinales en presencia del parásito41-42.
- Las interleuquinas 4 y 13 (IL-4 e IL-13) producidas por las células auxiliadoras tipo 2 innatas y por las células CD4+ Th2 modulan las funciones de los macrófagos circundantes de dos maneras: primero, deprimen las actividades proinflamatorias y microbicidas de los macrófagos de activación clásica (M1)7,43; segundo, promueven la secreción de las citoquinas antinflamatorias e inmunorreguladoras IL-10 y TGF-β por los macrófagos de activación alternativa (M2)7,44. IL-10 y TGF-β regulan respuestas inflamatorias e inmunitarias tanto de tipo 1 como de tipo 23.
Productos de excreción-secreción de helmintos estimulan cambios en la actividad de las células dendríticas.
La presencia de productos de excreción-secreción de helmintos modifica la actividad presentadora de antígenos de las células dendríticas de dos maneras7,25-26:
- Menor expresión en su superficie de moléculas coestimuladoras (CD40, CD80 y CD 86) y disminución de la síntesis de mediadores proinflamatorios, como IL-12 y TNF (del inglés tumor necrosis factor). Ambos hechos traen como consecuencia una supresión de la diferenciación de células Th0 a células Th1 o células Th175,7,45-49.
- Incremento de la producción de las moléculas antinflamatorias e inmunorreguladoras IL-10 y TGF-β. Esto, favorece indirectamente la diferenciación de células Th0 a células Th25,7,49-50.
Exposición de células dendríticas precursoras a helmintosque puede diferenciarlas a células dendríticas reguladoras, por factores no conocidos5 ,7,51-52 .
A diferencia de las células dendríticas clásicas, estas son ineficientes en la presentación antigénica a células T. En lugar de ello, estas células promueven la diferenciación de células Th0 nativas a células Treg o, alternativamente, estimulan la expansión de subpoblaciones de células Treg prexistentes.
Tomados de conjunto, los eventos descritos actúan en sinergia y el resultado es la modulación de las respuestas inmunitarias del hospedero, lo que compromete las respuestas Th1 y favorece la actividad de las células Treg7.
Cambios en la flora bacteriana del hospedero
La población de microorganismos normalmente presente en el intestino de los mamíferos es considerada su microbiota27. Observaciones diversas evidencian de manera creciente que esos microorganismos son importantes en la regulación de la homeostasis de sus hospederos y, consecuentemente, en el desarrollo de numerosas enfermedades53. Diferentes funciones fisiológicas, entre ellas las respuestas inmunitarias, el metabolismo y el desarrollo cognoscitivo, están afectadas por la composición de la microbiota54. Varios artículos de revisión sobre la relación entre microbiota y enfermedades humanas han sido publicados recientemente55-56.
Cambios en la flora bacteriana intestinal se han relacionado con las alteraciones inmunitarias presentes en enfermedades inflamatorias, tanto de localización intestinal como a nivel sistémico56-59.
Un desbalance en las poblaciones bacterianas comensales, conocido como disbiosis, es una característica común de los pacientes de enfermedades inflamatorias intestinales (colitis ulcerativa y enfermedad de Crohn)60.
Si bien la microbiota está restringida al tracto intestinal, los metabolitos producidos por las comunidades de bacterias pueden influir sobre órganos y tejidos lejanos. Un estudio demostró que la alimentación de ratones con una dieta rica en fibras conduce a un incremento en la circulación de ácidos grasos de cadena corta (AGCC), como resultado del procesamiento de las fibras por la microbiota intestinal61. Este incremento de los AGCCs regula la inflamación intestinal y promueve la diferenciación de células Treg Foxp3+62-63. Estos hallazgos podrían proveer una explicación a la contribución de algunos hábitos alimentarios de la vida moderna (por ejemplo, dietas ricas en grasas y pobres en fibras) al incremento de la susceptibilidad a enfermedades inflamatorias27.
Aunque los helmintos pueden modular directamente el sistema inmunitario de sus respectivos hospederos, tal como se ha descrito anteriormente, también estos parásitos pueden influir sobre las respuestas inmunitarias mediante sus efectos sobre la flora bacteriana intestinal22,64. Los resultados de trabajos recientes sugieren que la infección crónica por helmintos afecta la composición de la microbiota:
- La infección crónica por Heligmosomoides polygyrus favorece la presencia de determinados tipos de bacterias comensales, de manera particular miembros de la familia Lactobacillaceae65.
- La composición microbiana fecal de individuos que viven en países europeos industrializados es diferente de la de aquellos que habitan en áreas rurales africanas. Por ejemplo, el género Prevotella es más común en las personas que viven en países desarrollados 66. Esa diversidad bacteriana pudiera ser consecuencia de diferencias dietéticas, pero también de diferencias en la prevalencia de infecciones por helmintos64.
- Estudios muy recientes sugieren que los helmintos incrementan la producción de IL-22 por la mucosa intestinal, tanto en humanos como en ratones 64,67. La IL-22, además de estimular la producción de mucus y facilitar la reparación tisular, promueve la producción de péptidos antimicrobianos, incluidas proteínas de la familia de las β-defensinas68-73. A juicio de algunos autores, la IL-22 inducida por la infección por helmintos pudiera contribuir al control de la inflamación durante las enfermedades inflamatorias intestinales mediante la contención de la multiplicación de algunos tipos de bacterias22.
UTILIZACIÓN DE HELMINTOS EN EL TRATAMIENTO DE ENFERMEDADES HUMANAS
Evidencias epidemiológicas, clínicas e inmunológicas devenidas de estudios en humanos y datos obtenidos mediante experimentos en modelos animales ofrecen suficiente soporte al criterio de que las infecciones por helmintos tienen un efecto benefactor sobre entidades patológicas que transcurren con desregulación del sistema inmunitario, tales como alergias, enfermedades autoinmunes y alteraciones inflamatorias intestinales. Ese criterio condujo al desarrollo de numerosos ensayos de administración de helmintos vivos como herramienta terapéutica2,5,9,24,30-31,34 .
El primero de ellos data de 2003, cuando Summers y cols. reportaron el empleo exitoso de huevos de Trichuris suis en el tratamiento de enfermedades inflamatorias intestinales74. Desde entonces, además de los huevos de T. suis, también se han utilizado con fines terapéuticos el desarrollo de infecciones por Necator americanus. A continuación se describen los resultados de algunos de los trabajos en los que se ha empleado esta aproximación terapéutica en el control de enfermedades autoinmunes e inflamatorias:
- El T. suis es un parásito de cerdos que, aunque muy relacionado con T. trichiura, no puede mantener la infección crónica en los humanos y es eliminado en pocas semanas. Por ese motivo, el tratamiento con sus huevos consiste en la administración repetida de estos cada dos o tres semanas. En el estudio antes mencionado, el tratamiento resultó en una tendencia a la mejoría clínica tanto de los pacientes de enfermedad de Crohn como los de colitis ulcerativa74. Trabajos posteriores con pacientes de estas enfermedades que emplearon la misma herramienta terapéutica, pero realizados a mayor escala y con la utilización de grupos controles, confirmaron los primeros resultados 24,75-76.
- En pacientes de esclerosis múltiple, la administración de huevos de T. suis resultó en una tendencia hacia la reducción del número de lesiones nuevas, demostrada por imágenes de resonancia magnética77.
- Dado que T. suis es un parásito no adaptado a humanos, su administración está asociada al desarrollo de reacciones colaterales, fundamentalmente digestivas78. Esta adversidad ha conducido a algunos grupos de trabajo al empleo de parásitos de humanos con fines terapéuticos (por ejemplo, N. americanus). En pacientes de esclerosis múltiple, la infección por N. americanus resultó en una tendencia a la reducción de las manifestaciones clínicas asociadas a esta enfermedad79.
Sin embargo, los resultados de los estudios en los que se administraron helmintos vivos como herramienta terapéutica no siempre han sido favorables 5,22,24,34. Algunos, como los mencionados, demostraron la acción supresora de los parásitos suministrados 24,74-77,79. Otros, en cambio no encontraron el efecto protector esperado22,24,27,80-82.
Para entender esos resultados, aparentemente disímiles, deben tenerse en cuenta varias limitaciones presentes en parte de los ensayos hasta ahora realizados. Así:
1- Las personas que viven en áreas endémicas de helmintos están expuestas a infecciones parasitarias desde edades muy tempranas, generalmente antes de que tenga lugar el desarrollo de enfermedades con compromiso de la regulación del sistema inmunitario. Por ese motivo, la administración de helmintos con fines terapéuticos debe realizarse a edades tan tempranas como aquellas a las que ocurre la infección natural por esos parásitos. Sin embargo, debido a consideraciones éticas y prácticas muy comprensibles, la mayoría de los ensayos han sido llevados a cabo cuando los individuos participantes ya padecen las enfermedades que cursan con desregulación del sistema inmunitario y en consecuencia, cuando es más difícil suprimir desequilibrios ya establecidos5,22,83.
2- También por consideraciones éticas, muchos de los ensayos realizados han utilizado dosis parasitarias muy inferiores a las que ocurren durante la infección natural y podrían no haber sido eficaces en suprimir la desregulación inmunitaria de la enfermedad tratada84.
Además de estas limitaciones, otro importante obstáculo se erige frente a la utilización de helmintos vivos con fines terapéuticos: la no adherencia de parte de los pacientes al tratamiento porque este los haría portadores de parásitos vivos2.
Teniendo en cuenta estas dificultades, y las devenidas del empleo de helmintos vivos en el tratamiento de enfermedades alérgicas, los objetivos inmediatos son el logro de un conocimiento más preciso de los mecanismos por los cuales estos parásitos ejercen sus efectos protectores y, paralelamente, la identificación delas moléculas helmínticas capaces de replicar esas acciones; informes recientes dan cuenta de avances en este sentido85-86. Ello propiciaría el logro del objetivo final de este acercamiento al tratamiento de enfermedades alérgicas, autoinmunes e inflamatorias, el desarrollo de uno o más medicamentos que conserven la capacidad protectora de los parásitos vivos, entrañen mínimos riesgos para los pacientes, sean mayoritariamente aceptados por estos y puedan ser producidos con adecuados controles de calidad.
REFERENCIAS BIBLIOGRÁFICAS
1- Smits HH, YazdanbakhshM. Chronic helminth infections modulate allergen-specific immune responses: protection against development of allergic disorders? Ann Med 2007; 39:428-39.
2- McKay DM. The therapeutic helminth? TrendsParasitol. 2009 Mar;25(3):109-14. doi: 10.1016/j.pt.2008.11.008.
3- Moreau E, Chauvin A. Immunity against helminths: interactions with the host and the intercurrentinfections. J Biomed Biotech 2010;2010:428593.doi:10.1155/2010/428593.
4- Bruschi F, Araujo MI, Harnett W, Pinelli E. Allergy and Parasites. J Parasitol Res. 2013;2013:502562. doi: 10.1155/2013/502562.
5- McSorley HJ, Maizels RM. Helminth Infections and Host Immune Regulation. ClinMicrobiol Rev 2012; 25:585-608.
6- Rujeni N, Taylor D, MutapiF.HumanSchistosome Infection and Allergic Sensitisation. J Parasitol Res. 2012;2012:154743. doi:10.1155/2012/154743.
7- Salgame P, Yap GS, GauseWC. Effect of helminth-induced immunity on infections with microbial pathogens. Nature Immunol.2013;14:1118-26.
8- Weinstock JV. The possible link between de-worming and the emergence of immunological disease. J Lab Clin Med 2002; 139:334-8.
9- Weinstock JV. The worm returns. Nature 2012 Nov; 491(7423):183-5.doi:10.1038/491183a.
10- Fleming JO, Cook TD. Multiple sclerosis and the hygiene hypothesis. Neurology. 2006 Dec; 67(11):2085-6.
11- Mutapi F, Imai N, Nausch N, Bourke CD, Rujeni N, Mitchell KM, et al. Schistosome infection intensity is inversely related to auto-reactive antibody levels. PLoS One 2011, May;6(5):e19149. doi: 10.1371/journal.pone.0019149.
12- Correale J, Farez M. Association between parasite infection and immune responses in multiple sclerosis. Ann Neurol. 2007; 61:97-108.
13- Correale J, Farez M, Razzitte G. Helminth infections associated with multiple sclerosis induce regulatory B cells. Ann Neurol. 2008 Aug; 64(2):187-99.doi: 10.1002/ana.21438.
14- Pearson DJ, Taylor G. The influence of the nematode Syphaciaoblevata on adjuvant arthritis in the rat. Immunology.1975; 29:391-6.
15- Osada Y, Shimizu S, Kumagai T, Yamada S, Kanazawa T. Schistosoma mansoni infection reduces severity of collagen-induced arthritis via down-regulation of pro-inflammatory mediators. Int J Parasitol. 2009; 39:457-64.
16- Cooke A, Tonks P. Infection with Schistosoma mansoni prevents insulin dependent diabetes mellitus in non-obese diabetic mice. Parasite Immunol. 1999; 21:169-76.
17- La Flamme AC, Ruddenklau K, Backstrom BT. Schistosomiasis decreases central nervous system inflammation and alters the progression of experimental autoimmune encephalomyelitis. Infect Immun. 2003 Sep; 71(9):4996 -5004.
18- Sewell D. Immunomodulation of experimental autoimmune encephalomyelitis by helminth ova immunization. IntImmunol.2003;15:59-69.
19- Huibregtse IL, van Lent AU, van Deventer SJ. Immunopathogenesis of IBD: insufficient suppressor function in the gut? Gut. 2007; 56:584-92.
20- Johnston MJ, Wang A, Catarino ME, Ball L, Phan VC, MacDonald JA, et al. Extracts of the rat tapeworm, Hymenolepis diminuta, suppress macrophage activation in vitro and alleviate chemically induced colitis in mice. Infect Immun. 2010 Mar;78(3):1364-75. doi: 10.1128/IAI.01349-08.
21- Hunter MM, Wang A, Parhar KS, Johnston MJ, Van Rooijen N, Beck PL, et al. In vitro-derived alternatively activated macrophages reduce colonic inflammation in mice. Gastroenterol.2010 Apr; 138(4):1395-405.doi: 10.1053/j.gastro.2009.12.041.
22- MishraPK, Palma M, Bleich D, Loke P, GauseWC. Systemic impact of intestinal helminth infections. Mucosal Immunol.2014; 7:753-62.
23- Flamme LA. The platinum age of parasitology: harnessing the power of the parasite. Parasite Immunol. 2015; 37:275-6. doi: 10.1111/pim.12195.
24- Fleming JO, Weinstock JV. Clinical trials of helminth therapy in autoimmune diseases: rationale and findings. Parasite Immunol.2015; 6:277-92.doi: 10.1111/pim.12175.
25- Shepherd C, Navarro S, Wangchuk P, Wilson D, Daly NL, Loukas A. Identifying the immunomodulatory component of helminths. Parasite Immunol.2015; 6: 293-303.
26- Nutman T. Looking beyond the induction of Th2 responses to explain immunomodulation by helminths. Parasite Immunol. 2015; 6:303-13.doi: 10.1111/pim.12194.
27- Loke P, Lim YAL. Helminths and the microbiota: parts of the hygiene hypothesis. Parasite Immunol. 2015; 6:314-23.doi: 10.1111/pim.12193.
28- Ezenwa VO, Jolles AN. Epidemiology. Opposite effects of anthelmintic treatment on microbial infection at individual versus population scales. Science.2015 Jan; 347(6218):175-7.doi: 10.1126/science.1261714.
29- Rook GA. Hygiene Hypothesis and autoimmune diseases. Clin Rev Allergy Immunol. 2012 Feb;42(1):5-15. doi: 10.1007/s12016-011-8285-8.
30- Elliott DE, Weinstock JV. Helminth-host immunological interactions: prevention and control of immune-mediated diseases. Ann NY AcadSci.2012; 1247:83-96.
31- Finlay CM, Walsh KP, Mills KH. Induction of regulatory cells by helminth parasites: Exploitation for the treatment of inflammatory diseases. Immunol Rev. 2014 May;259(1):206-30. doi: 10.1111/imr.12164.
32- Girgis N, Gundra U, Loke P. Immune regulation during helminth Infections. PlosPathog.2013; 9(4):e1003250. doi: 10.1371/journal.ppat.1003250.
33- Blacwell AD, Tamayo M, Beheim B, Trumble B, Stieglitz J, Hooper P. Helminth infection, fecundity, and age of first pregnancy in women. Science.2015; 350:970-2.
34- Pallardo Fernández I. Enfermedades autoinmunes, tratamiento con Trichuris suis y otros helmintos. ArsPharm.2015; 56(2):65-75.
35- Saenz SA, Noti M, Artis D. Innate immune cell populations function as initiators and effectors in Th2 cytokine responses.Trends Immunol. 2010 Nov;31(11):407-13. doi: 10.1016/j.it.2010.09.001.
36- Massacand JC, Stettler RC, Meier R, Humphreys NE, Grencis RK, Marsland BJ, et al. Helminth products bypass the need for TSLP in Th2 immune responses by directly modulating dendritic cell function. Proc Natl Acad Sci U S A. 2009 Aug 18;106(33):13968-73. doi: 10.1073/pnas.0906367106.
37- Taylor BC, Zaph C, Troy AE, Du Y, Guild KJ, Comeau MR, et al. TSLP regulates intestinal immunity and inflammation in mouse models of helminth infection and colitis. J Exp Med. 2009 Mar 16;206(3):655-67. doi: 10.1084/jem.20081499
38- Harris N. Advances in helminth immunology: optimism for future vaccine design? Trends Parasitol. 2011 Jul;27(7):288-93. doi: 10.1016/j.pt.2011.03.010.
39- Fallon PG, Ballantyne SJ, Mangan NE, Barlow JL, Dasvarma A, Hewett DR, et al. Identification of an interleukin (IL)-25-dependent cell population that provides IL-4, IL-5, and IL-13 at the onset of helminth expulsion. J Exp Med. 2006 Apr; 203:1105-16.
40- Moro K, Yamada T, Tanabe M, Takeuchi T, Ikawa T, Kawamoto H, et al. Innate production of T(H)2 cytokines by adipose tissue-associated c-Kit(+)Sca-1(+) lymphoid cells. Nature. 2010 Jan;463(7280):540-4. doi: 10.1038/nature08636.
41- Neill DR, Wong SH, Bellosi A, Flynn RJ, Daly M, Langford TK, et al. Nuocytes represent a new innate effector leukocyte that mediates type-2 immunity. Nature. 2010 Apr 29;464(7293):1367-70. doi: 10.1038/nature08900.
42- Price AE, Liang HE, Sullivan BM, Reinhardt RL, Eisley CJ, Erle DJ, et al. Systemically dispersed innate IL-13-expressing cells in type 2 immunity. ProcNatlAcadSci U S A. 2010 Jun 22;107(25):11489-94. doi: 10.1073/pnas.1003988107.
43- Martinez F, Helming L, Gordon S. Alternative activation of macrophages: an immunologic functional perspective. Annu Rev Immunol. 2009; 27:451-83. doi: 10.1146/annurev.immunol.021908.132532.
44- Murray PJ, Wynn TA. Protective and pathogenic functions of macrophage subsets. Nat Rev Immunol. 2011; 11:723-37.
45- White RR, Artavanis-Tsakonas K. How helminths use excretory secretory fractions to modulate dendritic cells. Virulence.2012; 3:668-77.
46- Everts B, Hussaarts L, Driessen NN, Meevissen MH, Schramm G, van der Ham AJ, et al. Schistosome-derived omega-1 drives Th2 polarization by suppressing protein synthesis following internalization by the mannose receptor. J Exp Med. 2012 Sep; 209 (10):1753-67,S1.
47- Al-Riyami L, Harnett W. Immunomodulatory properties of ES-62, a phosphorylcholine-containing glycoprotein secreted by Acanthocheilonemaviteae. EndocrMetab Immune Disord Drug Targets. 2012; 12(1):45-52.
48- Klaver EJ, Kuijk LM, Laan LC, Kringel H, van Vliet SJ, Bouma G. Trichuris suis-induced modulation of human dendritic cell function is glycan-mediated. Int J Parasitol.2013; 43:191-200.
49- Carvalho L, Sun J, Kane C, Marshall F, Krawczyk C, Pearce EJ. Review series on helminths, immune modulation and the hygiene hypothesis: mechanisms underlying helminth modulation of dendritic cell function.Immunology. 2009 Jan;126(1):28-34. doi: 10.1111/j.1365-2567.2008.03008.x.
50- Everts B, Smits HH, Hokke CH, Yazdankbakhsh M. Sensing of helminth infections by dendritic cells via pattern recognition receptors and beyond: consequences for T helper 2 and regulatory T cell polarization. Eur J Immunol. 2010; 40:1525-37.
51- Steinfelder S, Andersen JF, Cannons JL, Feng CG, Joshi M, Dwyer D, et al. The major component in schistosome eggs responsible for conditioning dendritic cells for Th2 polarization is a T2 ribonuclease (omega-1). J Exp Med. 2009 Aug; 206(8):1681-90.doi: 10.1084/jem.20082462.
52- El Kasmi KC, Qualls JE, Pesce JT, Smith AM, Thompson RW, Henao-Tamayo M, et al. Toll-like receptor-induced arginase 1 in macrophages thwarts effective immunity against intracellular pathogens. Nat Immunol. 2008 Dec;9(12):1399-406. doi: 10.1038/ni.1671.
53- Cho I, Blaser MJ. The human microbiome: at the interface of health and disease. Nat Rev Genet. 2012; 13:260-70.
54- Cryan JF, Dinan TG. Mind-altering microorganisms: the impact of the gut microbiota on brain and behaviour. Nat Rev Neurosci.2012; 13:701-12.
55- Huttenhower C, Kostic AD, Xavier RJ. Inflammatory bowel disease as a model for translating the microbiome. Immunity.2014; 40:843-54.
56- Clemente JC, Ursell LK, Parfrey LW, Knight R. The impact of the gut microbiota on human health: an integrative view. Cell.2012; 148:1258-70.
57- Devkota S, Chang EB. Nutrition, microbiomes, and intestinal inflammation. CurrOpinGastroenterol. 2013; 29:603-7.
58- Strober W. Impact of the gut microbiome on mucosal inflammation. Trends Immunol. 2013; 34(9):423-30.doi: 10.1016/j.it.2013.07.001.
59- Brusca SB, Abramson SB, Scher JU. Microbiome and mucosal inflammation as extra-articular triggers for rheumatoidarthritis and autoimmunity. CurrOpinRheumatol.2013; 26:101-7.
60- Tamboli CP, Neut C, Desreumaux P, Colombel JF. Dysbiosis in inflammatory bowel disease. Gut. 2004; 53:1-4.
61- Trompette A, Gollwitzer ES, Yadava K, Sichelstiel AK, Sprenger N, Ngom-Bru C. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat Med. 2014; 20:159-66.
62- Arpaia N, Campbell C, Fan X, Dikiy S, van der Veeken J, deRoos P, et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature. 2013; 504:451-5.
63- Smith PM, Howitt MR, Panikov N, Michaud M, Gallini CA, Bohlooly YM, et al. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science. 2013; 341: 569-73.
64- Leung JM, Loke P. A role for IL-22 in the relationship between intestinal helminths, gut microbiota and mucosal immunity. Int J Parasitol.2013; 43:253-7.
65- Walk ST, Blum AM, Ewing SA, Weinstock JV, Young VB. Alteration of the murine gut microbiota during infection with the parasitic helminth Heligmosomoides polygyrus. Inflamm Bowel Dis. 2010; 16:1841-9.
66- De Filippo C, Cavalieri D, Di Paola M, Ramazzotti M, Poullet JB, Massart S, et al. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc Natl Acad Sci USA. 2010 Aug; 107(33):14691-6.doi: 10.1073/pnas.1005963107.
67- Broadhurst MJ, Leung JM, Kashyap V, McCune JM, Mahadevan U, McKerrow JH, et al. IL-22þ CD4þ T cells are associated with therapeutic trichuris trichiura infection in an ulcerative colitis patient. Sci Transl Med. 2010 Dec; 2(60):60-88.doi: 10.1126/scitranslmed.3001500.
68- Sonnenberg GF, Fouser LA, Artis, D. Border patrol: regulation of immunity, inflammation and tissue homeostasis at barrier surfaces by IL-22. Nat Immunol. 2011 May; 12:383-90.doi: 10.1038/ni.2025.
69- Turner JE, Stockinger B, Helmby H. IL-22 mediates goblet cell hyperplasia and worm expulsion in intestinal helminth infection. PLoS Pathog. 2013; 9: e1003698.doi: 10.1371/journal.ppat.1003698.
70- Wolk K, Witte E, Wallace E, Döcke WD, Kunz S, Asadullah K, et al. IL-22 regulates the expression of genes responsible for antimicrobial defense, cellular differentiation, and mobility in keratinocytes: a potential role in psoriasis. Eur J Immunol. 2006 May; 36: 1309-23.
71- Liang SC, Tan XY, Luxenberg DP, Karim R, Dunussi-Joannopoulos K, Collins M, et al. Interleukin (IL)-22 and IL-17 are coexpressed by Th17 cells and cooperatively enhance expression of antimicrobial peptides. J Exp Med. 2006 Oct; 203(10):2271-9.
72- Zenewicz LA, Flavell RA. Recent advances in IL-22 biology. IntImmunol.2011 Mar; 23(3):159-63. doi:10.1093/intimm/dxr001.
73- Zindl CL, Lai JF, Lee YK, Maynard CL, Harbour SN, Ouyang W, et al. IL-22-producing neutrophils contribute to antimicrobial defense and restitution of colonic epithelial integrity during colitis. ProcNatlAcadSci USA.2013 Jul; 110(31):12768-73.doi: 10.1073/pnas.1300318110.
74- Summers RW, Elliott DE, Qadir K, Urban JF Jr, Thompson R, Weinstock JV.Trichuris suis seems to be safe and possibly effective in the treatment of inflammatory bowel disease. Am J Gastroenterol. 2003; 98:2034-41.
75- Summers RW, Elliott DE, Urban JFJr, Thompson R, Weinstock JV. Trichurissuis therapy in Crohn's disease. Gut. 2005; 54:87-90.
76- Summers RW, Elliott DE, Urban JF, Thompson RA, Weinstock JV. Trichuris suis therapy for active ulcerative colitis: a randomized controlled trial. Gastroenterol.2005; 128:825-32.
77- Fleming J, Isaak A, Lee JE, Luzzio CC, Carrithers MD, Cook TD, et al. Probiotic helminth administration in relapsing remitting multiple sclerosis: a phase 1 study. MultScler. 2011 Jun;17(6):743-54. doi: 10.1177/1352458511398054.
78- Bager P,Kapel C, Roepstorff A, Thamsborg S, Arnved J, Rønborg S, et al. Symptoms after ingestion of pig whipworm Trichuris suis eggs in a randomized placebo-controlled double-blind clinical trial. PLoS One. 2011;6(8):e22346. doi: 10.1371/journal.pone.0022346.
79- Croese J, O'neil J, Masson J, Cooke S, Melrose W, Pritchard D, et al. A proof of concept study establishing Necatoramericanus in Crohn's patients and reservoir donors. Gut. 2006; 55(1):136 -7.
80- Blount D, Hooi D, Feary J, Venn A, Telford G, Brown A, et al. Immunologic profiles of persons recruited for a randomized, placebo-controlled clinical trial of hookworm infection. Am J Trop Med Hyg. 2009 Nov; 81(5):911-6.doi: 10.4269/ajtmh.2009.09-0237.
81- Feary J, Venn A, Brown A, Hooi D, Falcone FH, Mortimer K, et al. Safety of hookworm infection in individuals with measurable airway responsiveness: a randomized placebo-controlled feasibility study. ClinExp Allergy. 2009 Jul; 39(7):1060-8.doi: 10.1111/j.1365-2222.2009.03187.x.
82- Bager P,Arnved J, Rønborg S, Wohlfahrt J, Poulsen LK, Westergaard T, et al. Trichuris suis ova therapy for allergic rhinitis: a randomized, double-blind, placebo-controlled clinical trial. J Allergy Clin Immunol. 2010 Jan;125(1):123-30.e1-3. doi: 10.1016/j.jaci.2009.08.006.
83- Hepworth MR, Hamelmann E, Lucius R, Hartmann S. Looking into the future of Trichuris suis therapy. J Allergy Clin Immunol. 2010; 125:767-8.
84- Mortimer K, Brown A, Feary J, Jagger C, Lewis S, Antoniak M, et al. Dose-ranging study for trials of therapeutic infection with Necator americanus in humans. Am J Trop Med Hyg. 2006; 75:914-20.
85- Robinson MW, Donnelly S, Hutchinson AT. A family of helminth molecules that modulate innate cell responses via molecular mimicry of host antimicrobial peptides. PLoSPathog. 2011; 7: e1002042.
86- Robinson MW, Donnelly S, Dalton JP. Helminthdefence molecules-immunomodulators designed by parasites!. Front Microbiol.2013; 4: 296.
Recibido: marzo
4, 2016.
Aceptado:
junio 24, 2016.
Dr. Luis Fonte Galindo . Instituto de Medicina Tropical "Pedro Kourí" . Autopista Novia del Mediodía km. 61/2, La Lisa, Apartado postal 601, Marianao 13. La Habana, Cuba. Email: luisfonte@infomed.sld.cu
{kind=link}