ARTÍCULO DE REVISIÓN
La molécula CD28 y su función en la activación de células T
CD28 and its role in T cell activation
Dra. Danay Saavedra Hernández
Instituto de Ciencias Básicas y Preclínicas «Victoria de Girón». Universidad de Ciencias Médicas de La Habana, Cuba.
RESUMEN
La molécula CD28 es considerada uno de los receptores coestimuladores más importantes en las células T, necesaria para la completa activación celular. En los últimos años se ha acumulado suficiente evidencia sobre su participación en los mecanismos conocidos como señal 2 de activación celular o coestimuladora. De esta forma se convierte en un blanco atractivo para estrategias terapéuticas en enfermedades autoinmunes y trasplante de órganos. Esta revisión se concentra en los principales mecanismos por los cuales esta molécula participa en la completa activación de las células T.
Palabras clave: coestimulación, CD28, activación célula T.
ABSTRACT
CD28 is a very important correceptor among T cells and provide positive signals that promote and sustain T-cell responses. In the last few years, CD 28 has become an interesting target in grafts and autoimmune diseases. This review will focus on the mechanisms whereby CD28 allowing a complete T cell activation.
Keywords: costimulation, CD28, T cell signaling.
INTRODUCCIÓN
La molécula CD28 es uno de los receptores coestimuladores más importantes descritos en células T, esencial para la completa activación de estas células. 1, 2
En la actualidad se conoce que unido a la señal emitida tras el reconocimiento del antígeno específico (péptido presentado en el contexto del complejo principal de histocompatibilidad (MHC)) por el complejo del receptor de linfocito T (señal 1), a las células T les resulta imprescindible señales coestimuladoras adicionales (señal 2) para lograr la completa activación. Además necesitan el concurso de citocinas (señal 3) que consolidan el proceso de activación. 3
El término coestimulación habitualmente
describe la modificación del proceso de activación de la célula
T a partir de la interacción de receptores expresados en las células
T con sus
ligandos expresados en las membranas de las células dendríticas
(CD) u otras células presentadoras de antígenos (APC). 1,
3
Aunque la activación de las células T puede ocurrir con una potente señal de su receptor de linfocito T (TCR), la coligación con CD28 es necesaria en la mayoría de las respuestas a un péptido antigénico 4, 5 sin esta, generalmente se induce un estado de no respuesta, anergia o muerte celular 6. Consecuentemente las vías de coestimulación son atractivos blancos terapéuticos en enfermedades que se asocian con respuestas inmunes dañinas o aberrantes como por ejemplo: enfermedades autoinmunes, respuestas a alérgenos o trasplante de órganos. 7, 8. Este trabajo persigue el objetivo de realizar una revisión sobre la molécula CD28 y su participación en la activación de los linfocitos T.
La molécula CD28
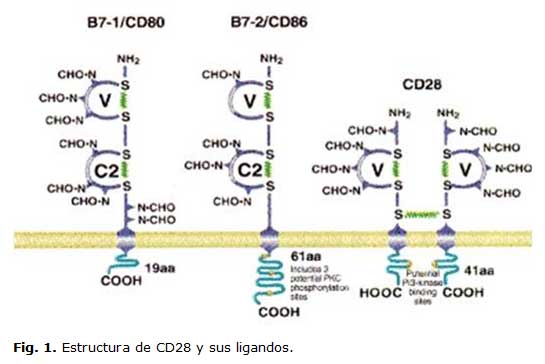
Esta molécula es una proteína de 44 kDa, miembro de la superfamilia de las inmunoglobulinas (Ig). Se expresa en la superficie de las células T como un homodímero estabilizado por un puente disulfuro. Sus genes están localizados en el cromosoma 2q33. 9
Estructuralmente forma un homodímero que se une a sus ligandos: CD80 (B7-1) y CD86 (B7-2), con una constante de disociación de aproximadamente 12 nM 10. (Figura 1)
Como molécula coestimuladora, sus principales funciones son:
Familia de B7:CD28
La familia B7:CD28 es el principal grupo de receptores/moléculas coestimuladoras involucradas en procesos de coestimulación y coinhibición en los linfocitos T. 13
Comprende los siguientes pares de receptores: ligandos:
La vía de señalización CD28/CTLA-4:CD80/CD86 es la mejor caracterizada. CD80 y CD86 comparten una especificidad dual para el receptor estimulador CD28 y para el inhibidor CTLA-4. 14
La familia CD28 de receptores se expresa por completo en los linfocitos T, el receptor PD-1 también se expresa en los linfocitos B y en las células mielocíticas. 13
Estos receptores son proteínas transmembranarias y todos incluyen un único dominio similar a la región variable de las Ig y una cola citoplasmática con tirosinas. 13
CD28, CTLA-4 e ICOS aparecen como homodímeros unidos por puentes disulfuro; PD-1 se expresa como monómero. 13
Vías de Señalización de CD28
El dominio citoplasmático de CD28, carece de actividad catalítica propia, por lo que el correceptor señaliza a través de la unión de proteínas señalizadoras intracitoplasmáticas. 15, 16
La activación de su vía de señalización, se asocia con la unión de varias proteínas señalizadoras a su motivo intracitoplasmático, formado por los residuos: Tirocina, Metionina, Asparagina, Metionina (TyrMetAsnMet o YMNM). En ocasiones la primera Metionina se sustituye por Valina (Tyr-Val-Asn-Met o YVNM). 12
La señalización se relaciona con
la composición de su pequeño dominio intracitoplasmático
de 44 aminoácidos y su interacción con proteínas intracelulares.
El motivo amino terminal (YMNM), una vez fosforilado en su residuo de tirosina,
se une a la subunidad p85 de PI3K (fosfatidilInositol 3 cinasa). 17, 18,
19, 20 Corriente abajo del motivo YMNM, la molécula CD28
cuenta con dos regiones ricas en prolina, (PRRP N terminal) que se unen a Itk
(cinasa inducible por IL-2). 21-22
La literatura consultada refiere la existencia de numerosas vías de señalización a partir de CD28, con la participación de diversas moléculas también involucradas en la transducción de señales de otros receptores como CD3, CD40, entre otros. 12, 23-25. Se detallan a continuación las tres vías mejor estudiadas y descritas.
Vía de la fostatidilinositol 3 cinasa (PI3K)
La unión de PI3K al motivo fosforilado de CD28 desencadena la producción de PIP2 (fosfatidilInositol bifosfato) y PIP3 (fosfatidilInositol trifosfato) que se unen a los dominios con homología pleckstrima en proteínas como PDK-1 (proteína cinasa dependiente de fosfoinositol 1), la cual activa la proteína cinasa B (PKB/AKT). Ambas, PDK-1 y PKB, pueden fosforilar otras proteínas y regular múltiples vías ligadas a síntesis de proteínas, metabolismo celular, y supervivencia. 23, 24 En este sentido, CD28PI3K y PKB proveen señales para aumento del metabolismo celular. 25
Está claro que la unión de CD28-YMNM fosforilado con PI3K genera señales pro supervivencia que previenen la apoptosis de la célula T. 26, 27, 28, 29
Vía de las Rac-MAP cinasas/Ras-Map-cinasas
El motivo YMNM puede igualmente acomodar la unión de la proteína adaptadora Grb2 (proteína 2 unidora del receptor de factor de crecimiento). La imposibilidad de unión de Grb2, por existir mutaciones en el residuo de asparagina (N), impide la fosforilación del factor de intercambio Vav1 (factor de intercambio de nucleótidos de guanina) y con este de la activación de JNK (serina/treonina cinasa c-Jun cinasa). 30 CD28 y Grb2 cooperan con Vav1 en la activación de los factores de transcripción NFAT (Factor nuclear) y AP-1(Proteína activadora 1). 31
Vav1 tiene un dominio (Dbl, dominio de homología) con actividad capaz de intercambiar GDP/GTP, para la activación de la proteína Rac1, la cual desencadena una cascada de activación de enzimas que culmina con la activación de la MAP cinasa JNK, 32 que propicia la síntesis y activación de factores de transcripción como AP-1.
Vav1 se une igualmente a PKCè (proteína cinasa C è), permitiendo igualmente la activación del factor de transcripción nuclear êB (NF-êB). 32
Vía del factor Nuclear Êappa B (NF-êB)
Han sido descritas otras vías mediante las cuales CD28 puede activar el factor de transcripción NF-êB. En este caso, su activación está regulada por una serina/treonina cinasa, IêB cinasa (IKKs), la cual promueve la fosforilación de IêB y su degradación por ubiquitinación, permitiendo al factor NF-êB entrar al núcleo. 33, 34, 35
La PKCè se transloca al sitio de unión de CD28 con su ligando, fosforila a la proteína Carma1 (caspasa con dominio de reclutamiento de membrana asociado a guanilato cinasa 1), la cual forma un complejo junto a Bcl10 y MALT1 (gen de translocación de linfoma de tejido linfoide asociado a mucosas 1). 36, 37
Este complejo recluta otra serina-cinasa, IKK, que al fosforilar a IêB (que se comporta como un inhibidor de NF-êB), es marcada para su degradación en el proteasoma. 38, 39
Al cesar la inhibición, NF-êB puede traslocarse al núcleo donde interviene en la transcripción de genes, por ejemplo de interleuquina 2 (IL-2). 38, 39
Principales resultados de estas vías de señalización
Coestimulación a través de CD28 en respuestas inmunes secundarias: rompiendo paradigmas
Estudios recientes en ratones han demostrado que es también útil recibir coestimulación para desarrollar una adecuada respuesta de linfocitos T de memoria. Investigadores de Filadelfia han observado que cuando la vía de coestimulación a través de CD28 se bloquea, ocurre una significativa reducción en la re-expansión de las células T CD8+ de memoria y en la magnitud de la respuesta que se genera contra el virus de la influenza. Se describe una reducción (de tres veces) en el número absoluto de células T CD8+ virus específicas, en ratones tratados con anticuerpos anti CD28, en comparación con los ratones del grupo control. Unido a este hallazgo se observó una significativa reducción en la función citolítica de estas células. 40
Las vías de coestimulación como dianas terapéuticas
Nuevos agentes terapéuticos han sido desarrollados para la supresión de respuestas inmunes dañinas sobre la base del conocimiento de estas vías de coestimulación antes mencionadas. 40
CTLA-4-Ig, medicamento biológico, consiste en el dominio extracelular de CTLA-4 y la porción Fc (Fracción cristalizable) de la IgG humana. Esta proteína se une a B7-1 y B7-2 y bloquea la interacción de CD28 con sus ligandos. Es conocido que la molécula CTLA-4 se une a sus ligandos con una afinidad entre 20 y 50 veces mayor que la afinidad mostrada por CD28 al unirse a estas mismas moléculas (ambos receptores comparten los mismos ligandos), es este el motivo por el cual no se ha usado la propia molécula CD28 para obtener producto. La unión de CTLA-4 a la porción de IgG, aumenta el tiempo de vida media de la proteína. 33
CTLA-4-Ig ha sido empleada en pacientes con Artritis Reumatoide, se desarrollan ensayos clínicos para evaluar su efectividad en el tratamiento del rechazo a trasplantes, psoriasis y enfermedad de Crohn. 40
En recientes investigaciones se ha determinado que las moléculas CD28 y CTLA-4 resultan atractivas dianas para el tratamiento contra algunos tipos de cáncer, entre ellos de mama y melanoma. 41, 42
REFERENCIAS BIBLIOGRÁFICAS
1. Acuto O, Mise-Omata S, Mangino G, Michel F. Molecular modifiers of T cell antigen receptor triggering threshold: the mechanism of CD28 costimulatory receptor. Immunol Rev 2003;192:2131.
2. Rudd CE, Taylor A, Schneider H. CD28 and CTLA-4 coreceptor expression and signal transduction. Immunol Rev 2009; 229:1226.
3. Leitnera J, Grabmeier-Pfistershammerb K, Steinbergera. Receptors and ligands implicated in human T cell costimulatory processes. Immunology Letters 2010; 128: 8997.
4. Bluestone J. New perspectives of CD28-B7 mediated T cell costimulation. Immunity 1995;2:5559.
5. Linsley PS. Distinct roles for CD28 and cytotoxic T lymphocyte-associated molecule 4 receptor during T-cell activation. J Exp Med 1995;182:28992.
6. Rudd CE. The reverse stop-signal model for CTLA4 function. Nat Rev Immunol 2008;8:15360.
7. Podojil JR, Miller SD. Molecular mechanisms of T-cell receptor and costimulatory molecule ligation/blockade in autoimmune disease therapy. Immunol Rev 2009;229:33755.
8. Li XC, Rothstein DM, Sayegh MH. Costimulatory pathways in transplantation: challenges and new developments. Immunol Rev 2009;229:27193.
9. Garçon F, Patton DT, Emery JL, Hirsch E, Rottapel R, Sasaki T, et al. CD28 provides T-cell costimulation and enhances PI3K activity at the immune synapse independently of its capacity to interact with the p85 D p110 heterodimer. Blood 2008;111:146471.
10. van der Merwe P, Bodian D, Daenke S, Linsley PS, Davis SJ. CD80 (B7-1) binds both CD28 and CTLA-4 with a low affinity and very fast kinetics. J Exp Med 1997;185:393403.
11. Chattopadhyay K, Lazar-Molnar E, Yan Q, Rubinstein R, Zhan C, Vigdorovich V, et al. Sequence, structure, function, immunity: structural genomics of costimulation. Immunol Rev 2009;229:35686.
12. Schneider H, Smith X, Liu H, Bismuth G, Rudd C. CTLA-4 expression disrupts ZAP-70 microcluster formation, T-cell D APC conjugation and calcium mobilization. Eur J Immunol 2007;38:407.
13. Gary A. Koretzky GA. T Lymphocyte Signaling Mechanisms and Activation. En: Paul W.E. Fundamental Immunology. 6th edition. Washington, DC: Lippincott Williams & Wilkins; 2008.347-72.
14. Driessens G, Kline J, Gajewski TF. Costimulatory and coinhibitory receptors in anti-tumor immunity. Immunol Rev 2009;229:12644.
15. Kölsch U, Arndt B, Reinhold D, Lindquist JA, Jüling N, Kliche S, et al. Normal T-cell development and immune functions in TRIM-deficient mice. Mol Cell Biol 2006;26:363948.
16. Wang H, Rudd CE. SKAP-55, SKAP-55- related and ADAP adaptors modulate integrin-mediated immune-cell adhesion.Trends Cell Biol 2008;18:48693.
17. Harada Y, Ohgai D, Watanabe R, Okano K, Koiwai O, Tanabe K, et al. A single amino acid alteration in cytoplasmic domain determines IL-2 promoter activation by ligation of CD28 but not inducible costimulator (ICOS). J Exp Med 2003;197:25762.
18. Tavano R, Gri G, Molon B, Marinari B, Rudd CE, Tuosto L, et al. CD28 and lipid rafts coordinate recruitment of Lck to the immunological synapse of human T lymphocytes. J Immunol 2004;173:53927.
19. Holdford AD, Green JM, Levin SD, Denny MF, Straus DB, Link PS, et al. Proline residues in CD28 and the Src homology (SH)3 domain of Lck are required for T cell costimulation. J Exp Med 1999;190:37584.
20. Sanchez-Valdepenas C, Martin AG, Ramakrishnan P, Wallach D, Fresno M. NFkappaB-inducing kinase is involved in the activation of the CD28 responsive element through phosphorylation of c-Rel and regulation of its transactivating activity. J Immunol 2006;176:466674.
21. Kim HH, Tharayil M, Rudd CE. Growth factor receptor-bound protein 2 SH2/SH3 domain binding to CD28 and its role in co-signaling. J BiolChem 1998;273:296301.
22. D'Souza-Schorey C, Chavrier P. ARF proteins: roles in membrane traffic and beyond. Nat Rev Mol Cell Biol 2006;7:34758.
23. Rudd CE, Schneider H. Unifying concepts in
CD28, ICOS and CTLA4 co-receptor signalling. Nat Rev Immunol 2003;3:54456
.
24. Alessi DR, Deak M, Casamayor A, Caudwell FB, Morrice N, Norman DG, et al. 3-Phosphoinositide-dependent proteinkinase-1 (PDK1): structural and functional homology with the Drosophila DSTPK61 kinase. CurrBiol 1997;7: 77689.
25. Frauwirth KA, Riley JL, Harris MH, Parry RV, Rathmell JC, Plas DR,, et al. The CD28 signaling pathway regulates glucose metabolism. Immunity 2002;16:769777.
26. Schneider H, Valk E, Leung R, Rudd CE. CTLA-4 activation of phosphatidylinositol 3-kinase (PI 3-K) and protein kinase B (PKB D Akt) sustains T-cell anergy without cell death. PLoS ONE 2008;3(12):e3842.
27. Kirchhoff S, Muller WW, Li-Weber M, Krammer
PH. Up-regulation of c-FLIPshort and reduction of activation-induced cell death
in CD28-costimulated human T cells. Eur J Immunol 2000;30:276574.
28. Jones RG, Elford AR, Parsons MJ, Wu
L, Krawczyk CM, Yeh WC, et al. CD28-dependent activation of protein kinase B
D Akt blocks Fas-mediated apoptosis by preventing death-inducing signaling complex
assembly. J Exp Med 2002;196:33548.
29. Wood JE, Schneider H, Rudd CE. TcR and TcR-CD28
engagement of protein kinase B (PKB D AKT) and glycogen synthase kinase-3 (GSK-3)
operates independently of guanine nucleotide exchange factor VAV-1. J BiolChem
2006;281:3238594 .
30. Kim H-H, Tharayil M, Rudd CE. Growth factor receptor-bound protein 2 SH2 /SH3 domain binding to CD28 and its role in cosignaling. J Biol Chem 1998;273:296301.
31. Schneider H, Rudd C. CD28 and Grb2, relative to Gads or Grap, preferentially co-operate with Vav1 in the activation of NFAT D AP-1 transcription. Biochem Biophys Res Commun 2008;369:61621.
32. Bustelo XR. Regulatory and signaling properties of the Vav family. Mol Cell Biol 2000; 20:146177.
33. Abbas A.K, Lichtman AH, Pillai S. Activation of T Lymphocytes. En: Abbas A.K, Lichtman AH, Pillai S. Celular and Molecular Immunology. 7th edition. Washington, DC: Elsevier Saunders;2011.203-24.
34. Hara H, Wada T, Bakal C, Kozieradzki I, Suzuki S, Suzuki N, et al. The MAGUK family protein CARD11 is essential for lymphocyte activation. Immunity 2003;18:76375.
35. Wang D, Matsumoto R, You Y, Che T, Lin XY, Gaffen SL, et al. CD3 D CD28 costimulationinduced NF-kappaB activation is mediated by recruitment of protein kinase C-, Bcl10, and IkappaB kinase beta to the immunological synapse through CARMA1. Mol Cell Biol 2004;24:16471.
36. Tanner MJ, Hanel W, Gaffen SL, Lin X. CARMA1 coiled-coil domain is involved in the oligomerization and subcellular localization of CARMA1 and is required for T cell receptor- induced NF-kappaB activation. J BiolChem 2007;282:171417.
37. Annibaldi A, Sajeva A, Muscolini M, Ciccosanti F, Corazzari M, Piacentini M, et al. CD28 ligation in the absence of TCR promotes RelA D NF-kappaB recruitment and trans-activation of the HIV-1 LTR. Eur J Immunol 2008;38: 144651.
38. Sathish JG, Johnson KG, LeRoy FG, Fuller KJ, Hallett MB, Brennan P, et al. Requirement for CD28 costimulation is lower in SHP-1-deficient T cells. Eur J Immunol 2001;31:364958.
39. Taylor A, Akdis M, Joss A, Akkoç T, Wenig R, Colonna M, et al. IL-10 inhibits CD28 and ICOS costimulations of T cells via src homology 2 domain-containing protein tyrosine phosphatase 1. J Allergy ClinImmunol 2007;120:7683.
40. Boesteanu A, Katsikis P. Memory T cells need CD28 costimulation to remember. SeminImmunol. 2009; 21(2): 6977.
41. Janakiram M, Abadi YM, Sparano JA, Zang X. T cell coinhibition and immunotherapy in human breast cancer. Discov Med. 2012;14(77):229-36.
42. Garbe C, Peris K, Hauschild A, Saiag P, Middleton M, Spatz A, et al. Diagnosis and treatment of melanoma. European consensus-based interdisciplinary guideline - Update 2012. Eur J Cancer. 2012;48(15):2375-90.
Recibido: 14 de octubre de 2012.
Aprobado: 26 de noviembre de 2012.
Dra. Danay Saavedra Hernández.
Dirección particular: Calle K # 15037 e/ 7ma y D. Altahabana, Boyeros,
La Habana.
Teléfono: 6461147
E-mail: danay.saavedra@infomed.sld.cu